 )
)
ͬһ��������è�����۾�����ȫ��ͬ�������Ƕ�����ɫä��������û�������ͬһ�뷹����è������ʲôζ����

�����ɶ�ϣ��óԲ��óԣ���
��ɲ���һ�����ĵ����⣬�����ζ���Ƿdz��ش����Ӧ�����⣬�ں�۽����ϰ����ž������صĵ�λ��ͬʱ������״�仯�ֵ����ط����ڷ��Ӳ��棬�������۽����ͺ�۽���������������
����ģ�������������ʼ�ͽ������ˡ�ζ�١���������ζ��ʶ��Ļ�����Ԫ���ֲ�����ͷ����������ʺ��ֲ��������������Ǹ����̳����ʴ����ͷ�Ϸֲ��� 9000 ���ζ�٣��ڲ��鶯��������һָ��
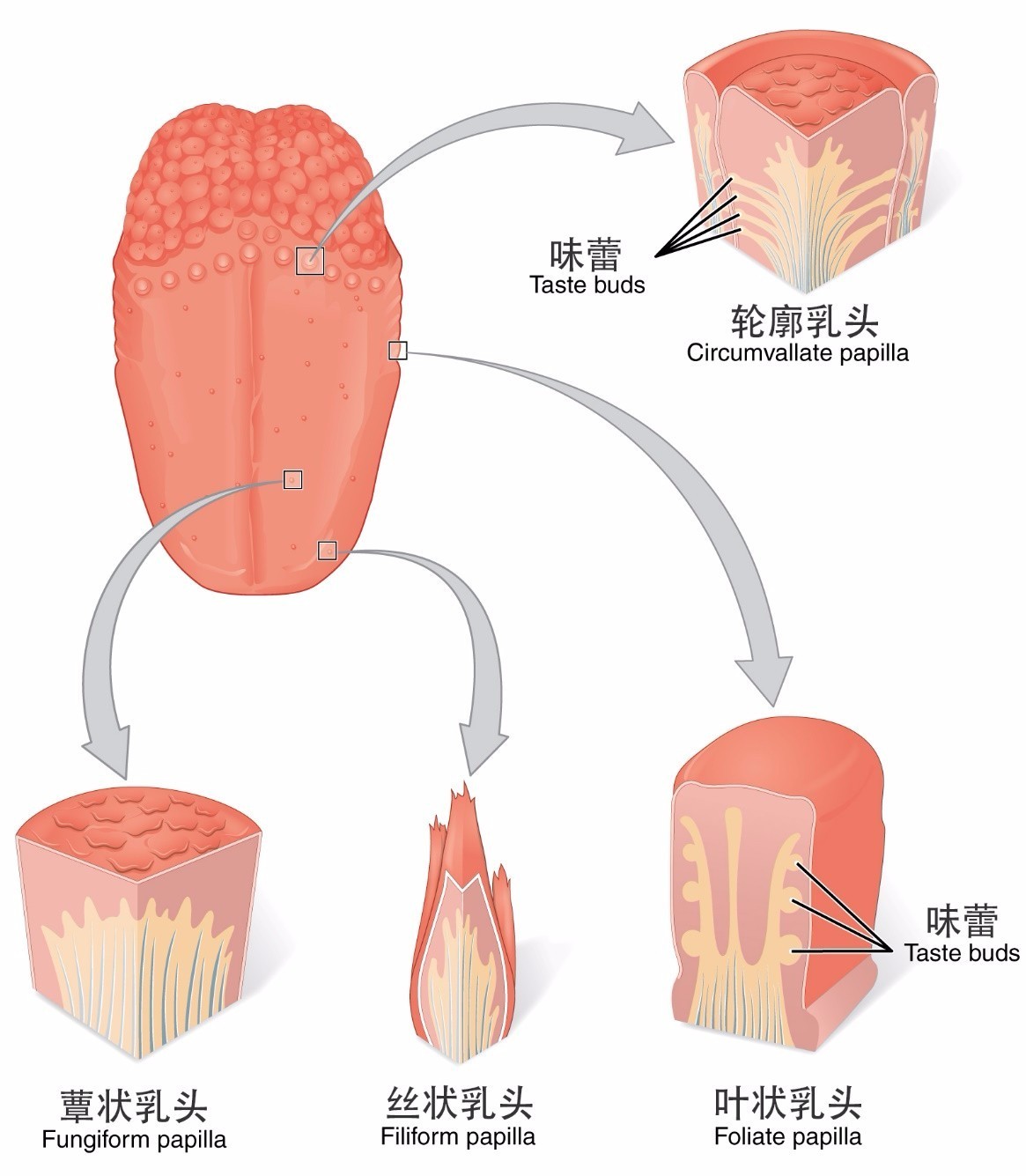
����Ϊ������ͷ�Ϸֲ������ֻ������͵���ͷ������˿״��ͷ���⣬��Χ����ζ�ٷֲ�
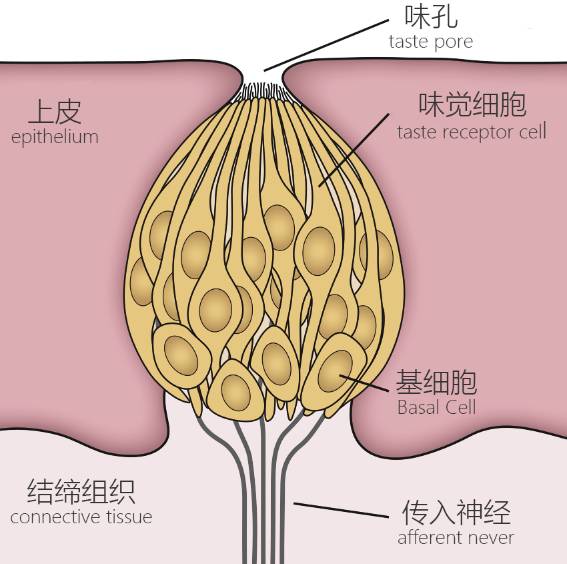
һ��������ζ��
ζ�ٷdz�����ͷ�����ɺܶ���״��ζ��ϸ��������ţ����ڿ�ǻ��Ƥ��dz��������ͨ��ζ�����ǻ��ͨ����Щζ��ϸ��������в�ͬ�����壬ÿһ�ֶ�ר�Ÿ���һ��ζ�����ۺ�������ÿ��ζ�ٶ��ֱܷ����е�ζ������һ���ģ����Ǹ��ݹ��ܽ�ζ��ϸ���ֳ������ࣺ
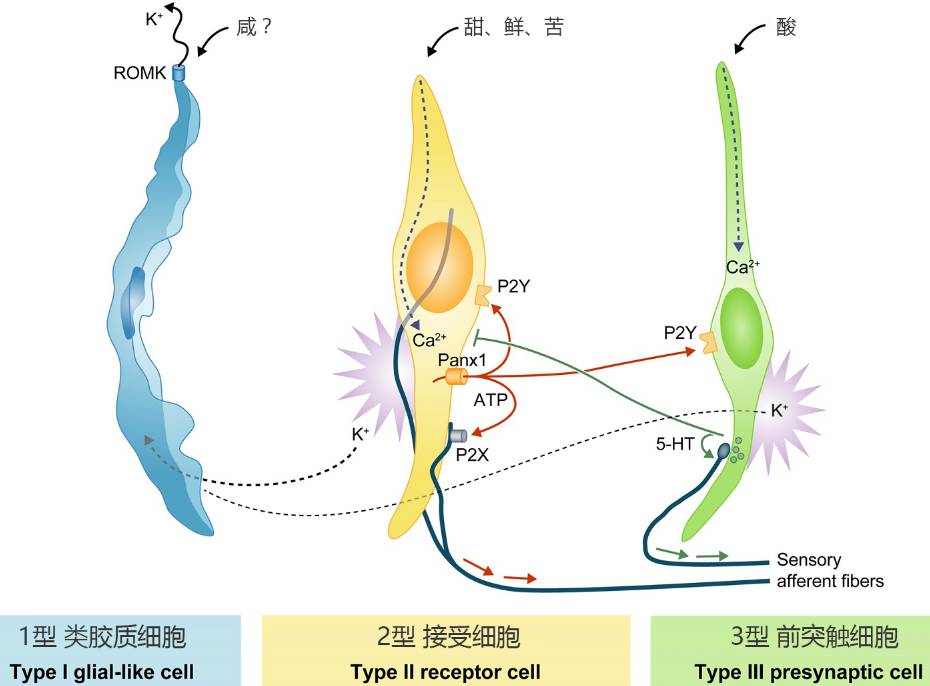
Ŀǰ�о�ȷ��������ζ������ϸ��
��һ���ɽ���ϸ���ػ��������DZ��溬�кܶ��Ƽ����ӱã������ų���������ϸ�����嶯���ͷŵļ����ӣ�ͬʱ����������ζ����Ҫ������������Ҳ����ζ�йأ�����������в����ˣ�
�ڶ����ǽ���ϸ�������ǵ�ϸ��Ĥ������Ƕ�˺ܶ� G ����ż�����壨G Protein-Coupled Receptors��GPCRs����������ϸ����౩¶�IJ����������Ե���ijЩ���ʽ�ϣ�ͬʱ��ϸ���ڲ⼤��һϵ���ź�ͨ·�����ոı�ϸ����״̬����������ͼ�Ľ���ϸ����������ϵ���ζ����ζ�Ϳ�ζ���ʺ����ջ�ó����� ATP��������һ��������λ��
�ڶ���ϸ����ǰͻ��ϸ�������DZ�����ϸ���ͷŵ�ATP������ͷų�5��ɫ�����ƽ���ϸ����ͬʱ��ˮ�������ӿ���ֱ��������ǰͻ��ϸ�������γɶ�����λ��������ζ��
��������ζ��ϸ���У�����ϸ������ؼ���һ�࣬��Ϊ��Ƕ��ϸ������� G ����ż�������൱��䣬���DZ�¶�IJ��־�����оһ����������Ϊ��ͬ�Ĺ��Ͷ�ƥ�䲻ͬ�����ʡ�����ζ���еĽ���ϸ����G ����ż������Ҳ�㷺�ֲ�����������������λ���Ǽ��ء����ﰷ���������ӡ����ʡ������������ζ���ӡ��������źŷ�����������壬������ 40%���ִ�ҩ�ﶼ�����ǵ����е㡣
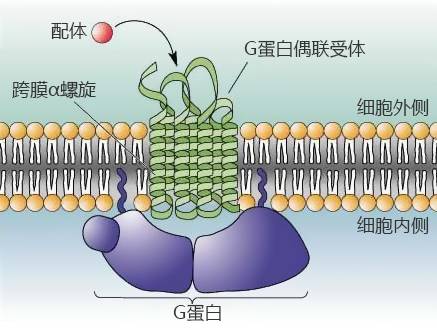
G ����ż��������Ƕ��ϸ��Ĥ�ϣ��ڲ��� G ������ϡ����������ϵ�ż�����屩¶�IJ��֣�ż������ͻᷢ��һ����̬�仯���������˶���ϸ���ڲ��� G ���ף�G ���ͻἤ��ϸ���ڵ��ź�ͨ·
G ����ż�������רһ�ԺͶ�������ζ�����Ǹ������ܻ���ͻ��Ӱ�죬���ոı䶯��ĺ����Ϊ�����������������ζ���ձ���������в��鶯���������⣬���ʺͿ��ζ�������� G ����ż������ı仯�ڲ�ͬ�ļ��������Ϸ���ǿ�ҵIJ��졪���ܵ���˵�������ܳ���ʲôζ���������ܳԵ�ʲô���ʸ߶��Ǻϡ�
����è��˵�����ǵ�ζ��������ķdz����ƣ����ڻ���ȶ��У����Ƿ������ǵ� TAS1R2 ������������� 247 �Լ�����������������������ζ�� G ����ż�����壬����è����ζ�����С�����ȷ��è�ƶ��ﶼ�ǵ�����ʳ���Թ�ʵ������Ȥ��Ҳȱ������ֲ����������������г����İ������֬����è�ƶ����ζ������������
����һ���棬èֻ�в��� 500 ��ζ�٣�������ͬ��ʳ����è�����û����ζҲ���ӹѵ����������Ǹ�������������ж�ʳ��óԲ��óԣ����������е�֬����ζ���������Ϊʲôè��������ζȴ��ϲ���Ա���ܣ��ⶫ�������������һ�������ζ�����ơ�

è��Ȼ��������ζ����������е����ͻӷ���������èҲ����ú�
������Ϊ��������ϵĻ�飬����ԭ���������Ҳ����ʳ�����������Ե�һ��ȥ�����ǵ�ζ��������û�����Բ�𣬵�ֻ�� 1500 ��ζ�٣�������èһ�������Ǹ��������ñ����ƶ�ʳ���ζ��������� G ����ż��������ࡣ
Ȼ��ζ�����˱���ʳ��ɷ֣����н�����ͷ���ʳ��Ϊ�Ĺ��ܣ�ij��ζ���źŶ�������˵�dz����ᣬ���Թ���˵���ܾ�������һ���¶���������������ʳ���ȱʱ�ܶ����ų� 6 ���֮���ʬ�壬����Ϊ�������Ѿ��ֽ�İ�����е��˷ܣ�ijЩ���������������ò����Ķ������Թ���˵ֻ�DZ��з�ζ������С�ԡ�

�����Լ������ˣ��������ӵ�ζ��δ���������������ô��
����è������������һЩ�dz�����Ķ���ζ������������ G ����ż�������йء��������ݶ����������Ƶij�Ա����һЩ�����ż�������ܸ��ܵ����۵�ζ�����������ǰ���ֲ��ĸ���������������ء������֮�£����������������ժȡ��ʵ����˶Ե��Ǻ�˫�Ǹ������У��Ե��۾ͺ���֪��

�����ȷ��ϲ�����ס��������ף��ȵȸ������۵�ʳ��
���������Ǵ���è��������ʳ��Ŀ�IJ��鶯����ȣ����� TAS1R1 ��������������λͻ�䣬ʧȥ�˶ԹȰ����ζ�������Ȱ���������ʳ��ζ����Ҫ��Դ�����Դ���è��Ϊʳ���ȴ�������⡪��������һ����ͻ���Լ������ 420 ����ǰ������ʯ֤�ݱ�������è�� 700 ����ǰ�Ϳ�ʼ�����ӣ�����ζ�Ŵ���è��ʳ��ת��Ҫ���ڻ���ͻ�䣬Ҳ����˵�ڳ��� 200 �������ʱ�������è�����ȶ����Ŷ���ʳ�Ŀ��������嵭�����ӡ�

����è��Ҫ������Ҷ������ĵ����ʺ����϶࣬���Բ������������ǵ��������������ڳ������кܶ���������ǰ���ά�طֽ���ǡ���������èӤ�����ò��ڳ�����֮ǰ�ȳԵ�ĸ�Ĵ�㣬�Ի����Щ������
����ζ�����ݻ���Ȼ�붯��ĺ����Ӧ���ܲ��ɷ֣���δ�ؾ�����Ӧ�Ե�ԭ������������Ӧ�ԵĽ�������鶯���ʳ�Ծ����൱�ߵĺ�������ԣ����ǿ�����Ϊ������ԭ���Բ����Ӵ�����ʳ�������ֳ�����������Ǵ���ʣ�������ζ��ͻ��ͻ��ý�����ͷ�ǿ������ѡ������������Ӧһ���µ���̬λ���������ݶ���ĵ���ζ�����������ͬʱ�����õ�ζ����ѻ��к�ͻ�䣬����ʧЧ��������è����ζ���ʹ���è����ζ��������
����پ�һ�����ҵ����ӣ��;�������й�һ���������ζ���ڲ��鶯������ٶۣ������� 3600 ����ǰ�� 5300 ����ǰ֮��ʧ�������������ζ��������������������ԵĿ�ζ�������������������ϰ�����̣�ȱ�������ϵĻ��ᣬ�ʺ��Ĵ���������������Ҫ�Ľ�ʳ�����������������Ӿ磬ʯ��й©�ͺ�ˮ��Ӫ�������ܲ��������ǿ�ҵĻ�����ʳ�Ǩ���еľ������ÿ�ζ�ֱ����е���в�����������վ������ڷ�����

2010 ���ī�������ʯ��й©�¼������˶�ͷĨ�㾨���������Ƕ�ʧ�˼�ʮ���Ϳ�ζ�йصĻ���֪�������ʳ�ﲻ�ܳԣ�Ҳ��֪����Щ�������Һ���ж�


������ ����Ķ�ԭ������������ʡ�































����˵�������а�