 )
)����01
����Բͷ����
����Բͷ����֢��һ�ֺ����ľ���ͷ��ȱ�ݣ�ϵ�Ծ��Ӷ����쳣Ϊ��Ҫ�����IJ����ۺ����������Բ��������з����ʣ�0.1%��4�ݡ��⾵�¾���ͷ����Բ�Ρ�����ṹ�쳣��ȱ�硢��������ܴ�������֮����1.00~1.12֮�䣬ͷ�����д��С�С֮�֡�����-��Ⱦɫ�У�ͷ����ʾ����ɫ����ʵ��ŨȾ��������ʾһϵ�еĵ���������ͼ3-ͼ4�����羵�¿ɼ�����ͷ��Բ������ṹ������Χ��һ����ȵĵ�λĤ����Ⱦɫ�ʾ���״��β�����������������Ҳ���ң�ͼ5����1971��Battaglia �����ȶ�Բͷ����֢����������������̬������Ҫ����������Σ����ز������������ȱ�磩���쳣��̬���Ӻ��Լ��쳣�ľ������������С�������֯��ѧ�о���ʾ��Բͷ����ȱ�����嵰�ף��磺����ø��������Ĥ��ԭ������ø���Ƽ���5�ݡ����߾�Һ��Բͷ�����������࣬��ʾ�䷢���������Ŵ����ػ������йء�
����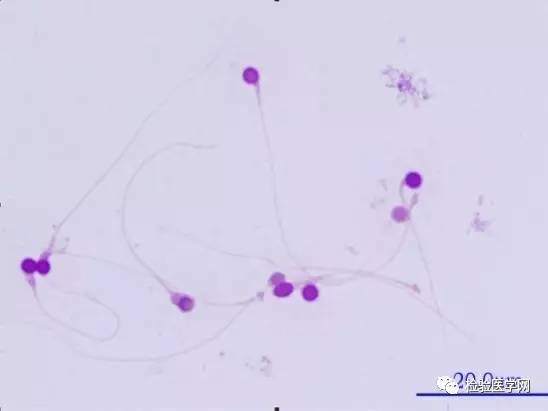
ͼ3Բͷ���ӣ�Բͷ������
����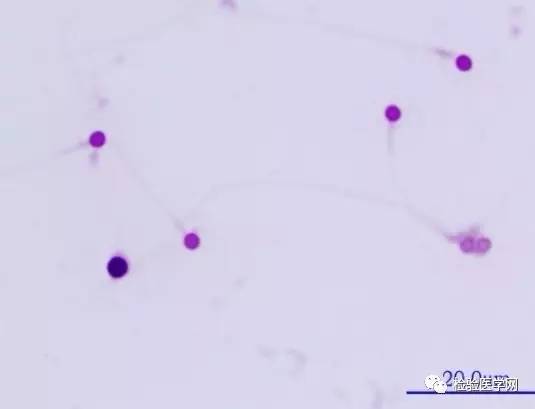
ͼ4Բͷ���ӣ�ͷ�˹�����
����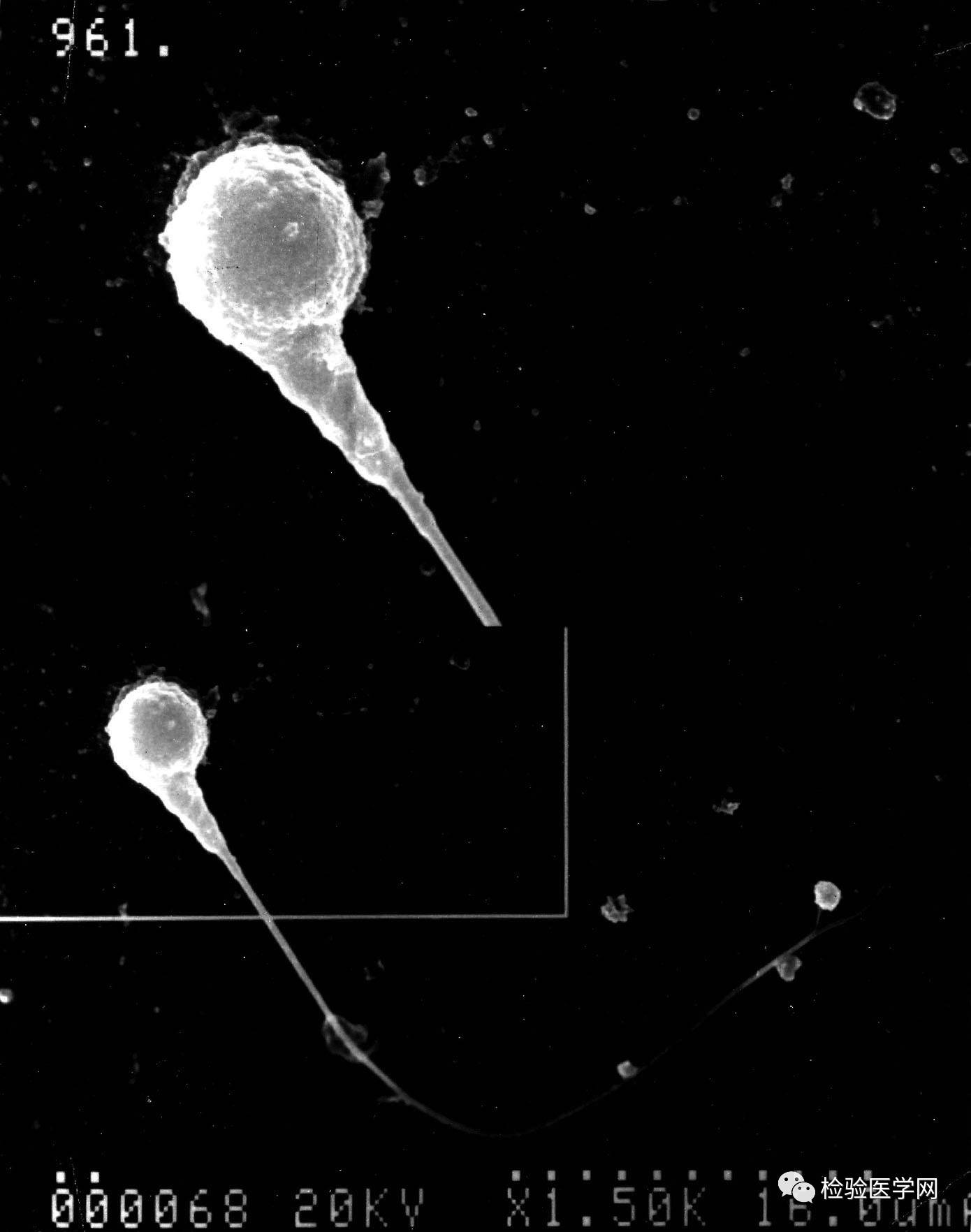
ͼ5Բͷ���ӣ�ɨ��羵��
����Singh ��6�ݽ�Բͷ���ӷ�Ϊ��������: ���ͽ�Ϊ���أ���ȫû�ж���Ͷ���ø; ���ͱ������ඥ�������������;�����̬�쳣��Kalahanis �ȣ�7���о���ʾ���������Ծ�Һ��Բͷ���ӱ�����2.3±0.5��%���������������ԣ�0.5±0.1��%����ʾ�������Ծ�Һ��Բͷ���ӱ������ӣ���������Ҳ���ӡ��о����֣�Բͷ����֢����������������ȣ�����ͷ�������ʡ�DNA ��Ƭ��һЩ�����Ե�Ⱦɫ��ķ������Ը������ԣ��ر���ijЩ�����ϸ�����ͻ���ȱʧ��
��������Բͷ����֢�ķ������Ƽ������в��������ȥ��Ҫ������Բͷ����֢����²�������о����������ӷ�����ػ���16( SPATA16) ������ø C1( PICK1) �� GOPC�� HIV1 ת����ϵ���( Hrb) ���ҵ���ø���2 �ǻ� ( Csnk2a2) �� bs ����ȡ������о�������һ�ֱ����Ĥ�ṹ�Ļ��� DPY19L2 ( dpy19like 2) �Ĵ���ȱʧ�ǵ��²������Բ���Բͷ���徫�ӵ���Ҫԭ�������µȣ�8���о������� DPY19L2 ����������λ���ͻ��Ӱ��һЩ���ذ������������ѧ���ʣ�ʹ�� DPY19L2 ���ײ��ܷ��������Ĺ��ܣ�Ӱ���˾��Ӷ�����γɡ������о�������9��10�ݣ����� DPY19L2 �ó��Ĵ�����ģ�ͣ�֤����ȱ�� DPY19L2 ����ᵼ�º����ܲ��Լ� acroplaxome �ͺ�Ĥ֮������Ӳ��ȶ���ʹ�ö����뾫��β�ܲ�����������ӣ������������ݲ����������䣬���Ӻ˲��ܳ��ͣ����վ��Ӷ��岻���γ�; �෴�ģ������������У������� DPY19L2 ����Ĵ���ȱʧ����Щ��֤ͬ���� DPY19L2 ����Ĵ���ȱʧ�ǵ���Բͷ����֢��һ����Ҫԭ�����������⣬�����ۼ����������(AR)���㾫����1����(PRM1)����-������ø 2 ����(GBA2)��˿���ᵰ��ø���Ƽ�����(PCI) �����������շ�ӦԪ�����������(CREM)�Ȼ�����Բͷ���ӵķ���������أ�11�ݡ�
��������Բͷ����ȱ������ø�����������ڻ����ⶼ������������ϸ���ܾ��������ICSI������������һ����Ψһ;���������ܾ��ʽϵ͡�
����02
���������
��������β���ṹ�쳣����ȱʧ�ǵ��¾��ӻ������µ���Ҫԭ��֮һ��ͼ6~ͼ7������Һ�о���100%���������ϲ���¼�������β���ӣ�ͨ�����������-������Ⱦɫ֤ʵΪ��������徫��ʱ����Ҫ���Ǿ��ӱ�ë�ṹȱ�ݵĿ��ܣ�ͨ����羵����ȷ��ϡ�Chemes �о����־������ƾ��ӱ�ë�ṹ�쳣��5������������Ϊ���صľ����˶��ϰ�����̬�쳣�����ṹ��ʾ������ά�ʼ���˿��װ�쳣������Ϊ��ά�ʷ�������(DFS)��12�ݡ�DFS ���߾�Һ�� 95% �� 100% ���Ӳ�����⾵��ɨ��羵�۲쾫�ӳʶ̡��ֺͲ�����β����羵�¼���ά�ʴ������ҵ�Χ���Ų�ͬ�̶ȱ��ε���˿���������������������ṹ������������ܼ��������ױ�ȱʧ;�ж�������ȱʧ�Լ�������ά����װ�쳣����ʾ DFS �Ǿ����γɹ������������ӱ�ë�ķ���������13��.
����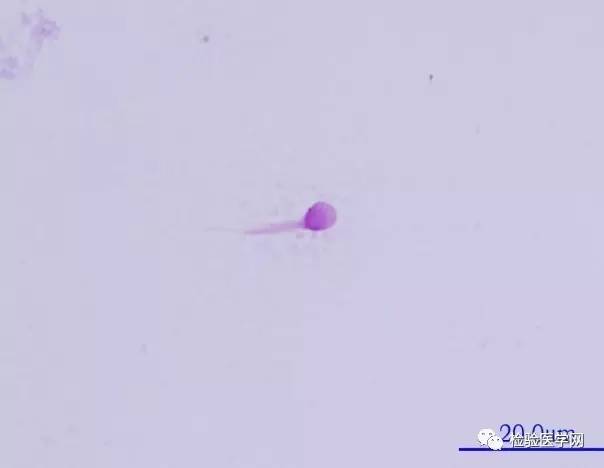
ͼ6��β����
����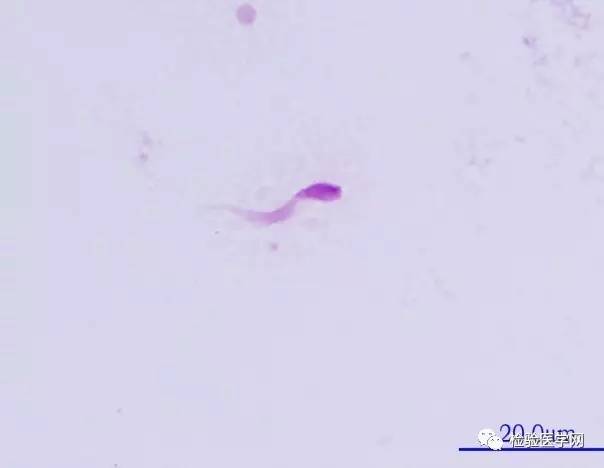
ͼ7��β����
����DFS�dz�Ⱦɫ�������Ŵ��������Ŵ�ѧ������δ��ȷ��������ʾ��14�ݣ���ϸ���ض�������������(Sptrx)��A ��øê������ 3 ����(AKAP3) �� A ��øê������ 4 ����(AKAP4)��ͻ���뾫����ά����װ�쳣������ء��������15���о����֣���β������ũҩ�Ӵ��ߵľ�Һ�г��ֱ��ʽϸߣ���ʾDFS���Ŵ�ѧ�����Ͽ��ܰ����������صĹ�ͬ���á�
����ԭ������ë�˶��ϰ���PCD������һ������������֢�ij�Ⱦɫ�������Ŵ�������DFS������Ϊ�����˶��ϰ�������ӡ�����������PCD�⾵�¾�����̬������������DFS����Ϊ��β���ӡ�ͬʱPCD�����з����ĺ���ϵͳ���䣬��֧�����ס����Ա���ǵ��ȡ��о����֣�16�ݣ�DNAJB13����ͻ����PCD�²����Ŵ�����֮һ��DNAJB13�Ա�ë�;���β����˿���γɺ��ܷ�����Ҫ�ĵ��ع��ܡ�
����DFS�������صľ����˶��ϰ�����̬�쳣��������Ȼ����ı��棬DFS ��Ӱ��ICSI���ܾ��ʺ��ٴ������ʣ����Ӵ��Ŵ�����ֵ�ù�ע��
����03
������ЧӦ����
��������غ��֮�������������⣬����Ϊ��ά�ֽϵ͵��¶ȣ��Ա�֤���ӵ����������ͳ��졣���Ҿ�������������չ���ص㣬�е��˵���غ���¶ȵ����Σ�ʹ��غ���¶ȱ���������¶ȵ�2�桫8�档���ҵ��¶ȵ��ڹ�������һ���ȵ�, ���¶ȵı仯�������ڷ�Χʱ , غ����������ܾͲ��ɱ�����ܵ�Ӱ�죬�������Ըı������Է�չ��
����غ���¶����߿ɵ��¾���Ũ���½����������ͼ����������ߡ��κ�ְҵ���ȱ�¶����������������������ϰ�ߵȶ���������غ���¶ȵ����ߣ����¶�Խ�ߣ�������ʱ��Խ������غ�����ֳ����Ӱ��Խ��ְҵ���ȷ��������¯�����纸������ʦ���������Ȼ����¹��������ȷ���Ӱ��ʹ�����¶����߽�������Һ�����½����ٴ��Ͼ����������ź���غ�Ļ����������½���������غ���¶�������һ���Ĺ�ϵ�����о��߱���, �����������Ż��ߴ���غ���¶��������� , ����Ϊ��������������غ�����µ�ֱ��ԭ��, ���¶�Խ�߾�Һ����Խ���غ������غ��δ�½������Ҳ������������ҵ��¶ȵ�����ϵ��ʹغ���¶����� , غ��������ϸ�������������ӣ�17�ݡ�����ԡ������Ȫ���������㡢��ʱ������Ȳ���������ϰ��Ҳ���������Ҿֲ��¶����ߣ�����Ӱ��غ�����ֳ���ܡ�
�������»�����غ���ڻ�������������л�������ĸı䣬���������ӣ�ø���Լ�ͨ�Ըı�ȣ����յ���֧��ϸ�����ԣ���������ϸ��Ӫ�����㡢�������˻����쳣���䣬�Ӷ�ʹ���Ӳ����ܵ����ơ�������������ϸ�����˳�����֧��ϸ����̬�빦�ܸı䡣��ЧӦ�����˾���DNA��RNA�͵����ʵĺϳɲ��������ʱ��ԡ�Ⱦɫ���װ�쳣��DNA���������ƻ����о���ʾ������ϸ����������ǵķ��������ճ�Ϊ����DNA������л����ľ��ӡ�˵����ЧӦ����DNA�������ԡ�غ���¶����ߣ��ɳ�����Դ�Գ�ͷ���ӣ����ص��Ǿ��Ӿ����쳤��ͷ���뾱�ж�ŨȾ��β����ȱ����������״̬��18�ݣ�ͼ8~ͼ9���������߾�Һ�г��ִ�����ЧӦ����ʱ��һ��Ҫ���Ǹ������صĴ��ڡ�
����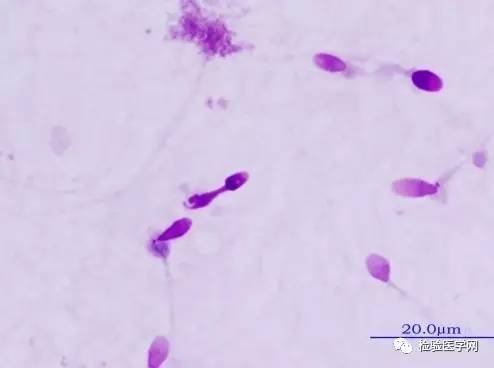
ͼ8��ЧӦ����
����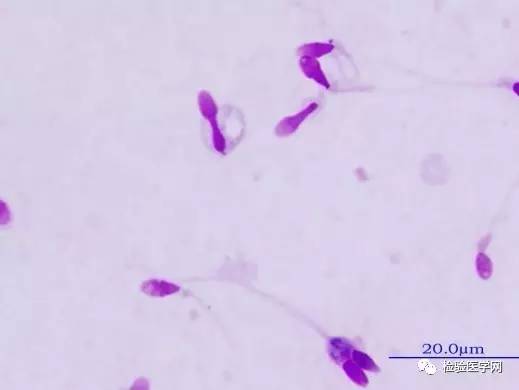
ͼ9��ЧӦ����
����04
������ͷ����
������ͷ�����������������Ծ�Һ���� ,һ��< 10% ,ͨ���ڲ������Ծ�Һ�б���Ϊ 2 %�� 20%[19]����ͷ�����������Ǿ��ӻ��װ�ǰû��Ⱦɫ�ʻ�ͷ���ṹ, �ھ�Һ�н�������β������ͷ��״ȱ�ݣ�[20](ͼ10~11)���߱�����ͷ���ӽ�����Ӱ�쾫Һ���� ,ʹ�����ܾ������½� ,���²�����
�������ִ�ͷ��״��������̬����ʱ����������ͷ��ȱ�ݽ��м�������Ӧ��Ϊ����ṹȱ�ݽ��б��桢��������ȻijЩ��Ϊ���������ĵȿ�ʹ����ͷβ����, ��������ת��<2 000 r/min, ����ʱ��<10 min, һ�㲻�����ͷβ���룬��Ϊ���е�����ز�����������������Ӷ��쳣, ͷβ���Ŵ�������, ��Һ�г��ִ�����ͷ���ӵ�ͬʱ,Ҳ����ִ���ȱβ���ӡ���ͷ����֢�ķ��������ͷβ���ӶεĻ���ȱ���йء����о���ʾ[21] ����������ά����1����ODF1���������������ʺ;���β������������ά����ȷ����������, ͬʱ���Ǿ���ͷβ�������������衣��״ͬϵ�ﵰ�� 1 ����( HOOK1)�Ծ��Ӿ��������ëϸ���ṹ����������Ҫ����, �书��ȱʧ����ɾ���ϸ�����ܽṹ��λ, ���¾���ͷβ�����쳣[22]�������о�֤ʵ[23]������ϼ�����о��ŶӶ�2�����Բ���֢���߽�����ȫ�����Ӳ�������SUN5�������1������ͻ��p.Thr275Met��2�������Ӻ�ͻ��p.Arg356Cys��p.Met162Lys������15�����Բ���֢���ߵ�SUN5�������Sanger��������������6�������з�����2������ͻ���4�������Ӻ�ͻ�䡣��ˣ� SUN5����ͻ�䵼�»��߳��쾫���иõ��ױ�����������������ȱʧ����Ӱ��SUN5�����ھ���ͷ����������ı��ﶨλ����ͻ���ǵ���������ͷ����֢����Ҫ�²�ԭ������������, ����SPEM1��GAT1��PRSS21 �Ȼ����뾫��ͷβ�������[24]��
������溱���[24]������ͷ���ӱ���Ϊ98%��20%��15%�Ļ��߽��о��Ӻ�DNA�����Լ�⣬�����ʾ��SDFI ֵ�ֱ�Ϊ95%��34%��27%��˵�����Ӻ�DNA���˳̶�����ͷ���ӱ�����������ߡ�����SDFIֵ�뾫�ӻ������нϸ������, �ر���SDFI>30%ʱ, �������������Խ��͡���Ȼ������ֳ����Ϊ������ṩ����������, ���ܾ��ʼ�Զ�ڰ�ȫ������֪̽��
����
ͼ10��ͷ����
����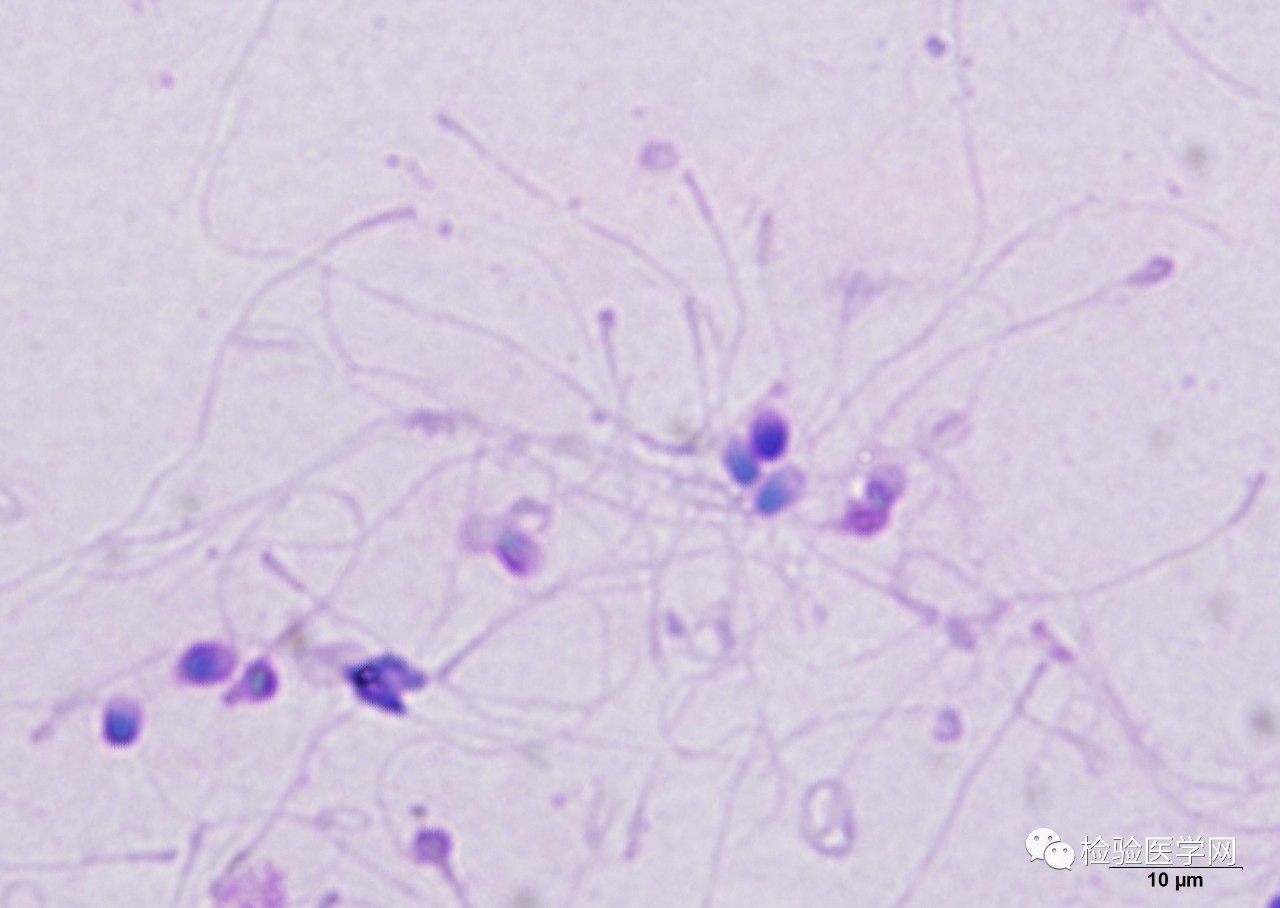
ͼ11������ͷ����
��������Ķ���
��������������������̬ѧ�ļ���Ŀ������״
�������쾫�ʼ����������˽���ָ����ľ�����̬�������ע��






























����˵�������а�