 )
)����3����Դ������ע "������" �Ź��ںţ��ظ��ؼ���"3��"����ȡDNA���з���������
Discussionд��ģ��| SCI��ͼ| qPCR����| �������mTOR�ź�| ELISAʵ��
����
�����༭���ӷ��㣨ת����ע��������·ҽ���������֣�
������ȥ��ŵ��лĻ֮��������Ϊϸ���ڵ������ά��ϸ������̬�����Ϊ��֪��һ��������Ϊ����������ְ��������������Ա�֤���ϸ��������ࡣ�����ƣ���ѹ�Ⱦͻ��з�������ѹ֮�£�����Ҳ��¶�����������ݣ��������������Լ����ȱ���������
����Science���ڵ�һƪ������ѹ��֮�£�����������;������·�̣��ź�ͨ·�����͵���������������ڸ�λ�����ߵİ����£�������λ�����ܹ����ձ��������졣
������˵��ϸ������Ի�ƽ����ϲ���Լ����Լ��棬ͨ�������ǻ��������ɣ�basal autophagy��ÿСʱ�ɷֽ��ϸ����1%��1.5%�Ĵ�л���ʡ����䵱ѹ������ʱ������������Ӫ����������лʧ�⡢ȱ��������Ӧ�����°���������ʱ��������ص��ף�ATG���ͻ�ߣ�����ӣ�ʹ����£��·ת¼������P53��FOXO��MiT/TFE��Nrf2��NFkB/Rel�ȣ�������������ģʽ����ϣ����ѹ��Ϊʳ��������֮�����ɷ�Ӧ��ϸ��Ӧ��Ӧ����Ӧ��һ������������
����???��ϸ���Ի�������
�������ɵ��γ�????

��������ˣ�������ȿ���ϸ������Ի�����ô���ɵİɣ�����ͼ����
������Ȼ����mTORC1���ڷǻ״̬ʱ��������ʼ������ķ�ӡ�������ʼ�������Ǩ������������Ĥ������ATG14��VMP1���ף���
�������ţ�ATG14��ULK1/2�����������ữ������ļBeclin1�����Լ�Vps34/15���ף����γ���֬��������ø3��PI3K��С�Ŷӣ�ר����ʹ��֬���������ữ��PI3P���Ļ����������ATG9�����Ե�����С����С�ܡ�
����ͬʱ��һЩ�����γ�������Ӳ����ATG12-ATG5-ATG16��LC3-PE������Ҳ��PI3PĽ��������ԭ����LC3-PE����ATG12-ATG5-ATG16&PI3K����һ����ֻͼ�������²���������������֮���ĬĬ��ʧ�����������ϣ���һ��ֻ��λ������Ĥ������ʵ�LC3-PE���������ˣ�LC3�ᱻ��ʱ������������dz�����ʵ�LC3-PE���ڶ�ػ����ۺ����p62�����¶���������Ĥ�ϡ�������ĤҲ��p62��������������첢���Ϊһ���������ݣ�����������ø���ɽ��ⱻ����������Ա���һ�١�
������ѹ֮��
�������ɵ��ɱ�

����������֪��mTORC1�������ź�ͨ·�������쵱���ĺ�������ⲻ֪��Ϊ��ϸ��������ҵ���������������Σ������������Ӵ̼���DNA���ˡ�����״̬��ATP/AMP��ֵ����������Ũ�ȣ�ͨ�����ӱ�V-ATPase�������������ɻ���ROS��Ũ�ȵȼ��ͳͳ���������ƿ�֮�С����ܴ����ʱ��������ͨ�����ữ���������ٽ�����ϳɣ����Ƕ������ɣ���ƫƫҪ���������֮����״̬�����ữ������ULK1/2��ATG13�����ԣ�����ʹ���������Ա�������
��������Ϊ��ά���������״̬��������ҪRheb�����Լ���λ����ø���Rag���ף�GTPase���Ķ��������¡�����Rag������ļ��TSC1/2���ף�GTPase����ף�GAP��ǡ������mTORC1�����һλ����ұۡ�����Rheb����GTP�����ʽ��ά�֣���mTORC1�̬�Ƿdz���Ҫ�ģ�һ����Rheb-GTP��ʽ�����ữ��TSC1/2תΪRheb-GDP��ʽʱ��mTORC1Ҳ���ڱ��ͷŵ�ͬʱʧȥ����ԣ������������ɷ�Ӧ��
������ʱ���Ͳ��ò��ἰ�����ε����ֵ���AKT��AMPK��AMPK�����Ǽ������������Ǵ�����̼��ź�����ԭȱ����AMP/ATP��ֵ���ߣ�����Ũ��ROS��DNA���ˣ�DDR������ʹ����������ữTSC1/2����������ģʽ���ɼ������ɷ�Ӧ��Ӧ�Ը���ѹ��Դ�Ĺ����д��ں��ĵ�λ������ǡ����Ϊ�����κ�ϸ�������ϰ�����л��ƽ����ᵼ���������ͷ�ROS�����������ɷ�Ӧ��
�������ɵ�����ת¼����֮
�����������
����1.ATG7&ATG5��Ӧ����Ӧ�ж�p53ͨ·��Ӱ��
����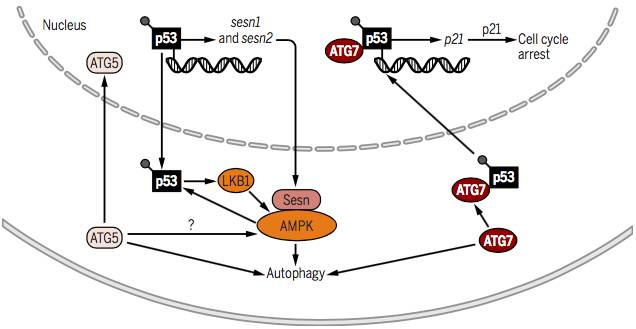
����ͨ��AMPK���������Ƶ���p53�����ε��ס���p53֮���������е�Ӧ����Ӧ�ж��ܷ�����Ҫ���ã�����������Ϊ����ʽ�����й�ϸ����л�����ɺ͵�����������Ҫ��ת¼���ӻ���Ҳ����Ϊ���ܳ�Ϊϸ�����е�ЧӦ���ӡ�
����ѹ��Ӧ����p53��Sesn1��Sesn2����ķ�ʽ����ɼ���AMPK���ٽ����ɷ�Ӧ��ͬʱ����ATG7�����£�p53��ʽ����ת¼����p21CDKN1A�ı����������G1��G2ϸ������ͣ�͡� ��ATG5Ҳ������p53��ȣ����ữ�������p21CDKN1A��ʽ������⣬������Ҳ���������ữp53���ڼ���LKB1�ɴ�ʹAMPK�������λ����ATG7��ATG5Ҳ�����յ����ɡ�
����
2. ATG7 �鵼FOXO1�ĸ���������
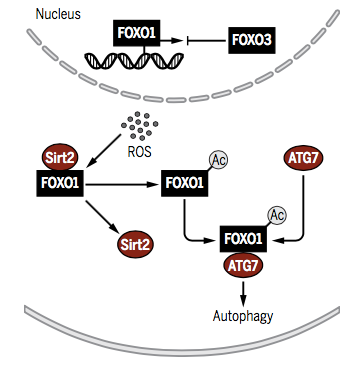
FOXOת¼������ѹ�������¿ɼ�������������ɵ���Ҫ����ת¼����FOXO3����FOXO1��ϸ�����еı���Ӷ��������ɡ�����������Ӧ����ROS�������������øSirt2ʧ���������FOXO1��FOXO1-Ac���������ϸ�����в���ATG7���ϣ������ɴ̼����ɷ��������������յ�������ʹ��FOXO1��ϸ�����б���ת¼����ʧ���״̬���Ӷ������������е�����������ֲ�����������⣬ATG7�ڷ��ػ������Ӧ�еļ�����γ�ATG5-ATG12�������LC3-��֬���Ҵ�����LC3-PE�����������������γɺͱպ�������Ҫ��
����3. p62������ѡ����������Ӧ����Ӧ;���в���Nrf2�����ķ�����
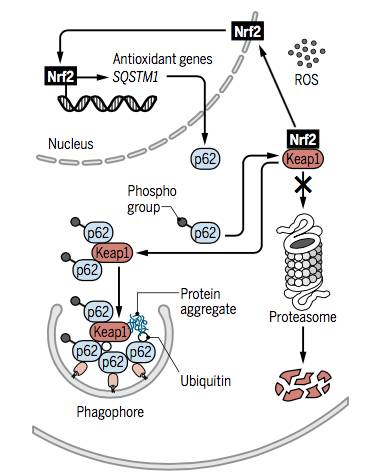
ѡ�������ɵĻ�����������p62����ص���������/�νӵ��������ԣ����У�p62�������幦��ʧ����ROS�������йأ�����Ӧ����Ӧ�п��������á����⣬p62Ҳ��һ����Ҫͨ��������������ĵ������һ��p62�����ͻ��յ�ROS������DNA���ˡ��������������������
������ˣ�Ϊ��ά��ϸ����������ԭƽ�⣬���ɹ�����Ҳ������һ��ǰ��;������ROSŨ������ʱ������p62��SQSTM1�������ת¼����Nrf2��ʽ��������忹�����ͽⶾ��Ӧ��������Ҫ���á����⣬p62Ҳ�ɵ���Nrf2�ķ�ȣ�������������øKeap1����Nrf2��ᱻ26S����ø�彵�⣻��p62����Keap1��Ϻɽ������������ػ������ʵľۼ���һ�������������е�����Ĥ�����ڿ��ٽ��⡣���⣬Ҳͨ��SESN2��ϼ�������ữ��p62��Keap1���������ӣ��Ӷ�ͨ������Keap1������������ǿ����������
����4.����������ʱ��ת¼����MiT / TFE�ĵ���ȡ����ATG5/9����
����
����������ʱ�������˶����ɺ���ø�嵰��������Щ������ı���ȡ����ת¼���ӵ�MiT/TFE���壨��TFEBΪ����������ͨ������mTORC1���ữ�������14-3-3������ϸ�����С��������幦���ϰ�������ROS���ɣ���øPINK1���ữE3��������øparkin������ļ������������Ĥ�������ػ�������������浰�ף�ʹ����Ϊ��������/��������p62��optineurin���İб꣬�����γ���������С�塣
����TFEB�������λ����ϸ����ȡ����parkin��ATG5��ATG9������ã����ܷ�ʽ����������ø������ɻ�������֤�ݱ�������ATGҲ����ϸ���ˣ�ATG5��ATG9�ڴٳ�����С���γɹ����и����ܵ������Ǵ�ʹTFEB��λ��ˡ����⣬�����ǰ���Ҳ�ɴ̼�TFEB�����λ������������ATG5��ATG9���ס�
����5.���ɺ�NFkB / IKK������������

����NFkBȷʵ�ɷ�ʽ��������������ػ�����Beclin 1 ���������NFkB��ص���֢��Ӧ�ź�ͨ·�������ź�ͨ·�����ص��ġ�
����ͨ�������е�NFkB�������Ƽ�IkB��ϣ�������IKK�����D��û�����IkB�ᱻ����ø�彵�⣬�Ӷ��NFkB��ʹ����λ����ϸ���ˡ���IKK�Ļ��ҪTGF-b���ø1��TAK1�����丨����TAB2��TAB3������TAB2��TAB3Ҳ����Beclin 1�γɸ��������������Bcl-2Ҳ����Beclin 1��ϣ�������NFkB����ֻ������Щƽ�ⷢ���仯�Ľ�����һ������ɷ�Ӧƽ�з���������TAB2��TAB3Ӧ���� IKK�����ͬʱ���ͷ�Beclin 1�������ɡ�
����6.UVRAG����Ϊ�������ɺͰ�֢��ͬ���ܵ���Ŧ
�������ڻ��۶��Ե����ʾۼ��壬����ʧ�����������ROS��������ȱ�ݿɵ���ϸ���еķ���������°�ת����������Ҳ��ͨ�������߷��俹����ػ���UVRAG�������Եر��������鲻�ȶ��ԡ�
����UVRAG�ɲ���Beclin 1��Vps34�γɸ�����PI3K�����������������γɡ���UVRAG����һ�����Ϳ��Խ��DNA�����Ե���ø��ʶ��DNA����λ���NHEJ�Ĺؼ�ø����������������ı��������⣬UVRAG������������ʼ�з������ã����ɵ����������������ڡ�������С�����ø���ں��У���mTORC1���ữUVRAG����ֹ��һ������������������ø����ںϣ������������ɽ������ʡ�
������֮����������Ӧ����Ӧ������ܽ�ϵ�һ��;������ATG����ȷ��������Ӧ�������е��ں���������ת¼���ӵĻ��ԡ�
��������Beclin 1�ɾ����Եؽ���漰NFkB��ĸ��ֵ����ʣ�����Ϊϸ������������֮���ƽ��㣻ATG5����p53��Ⱥͻ����������ת¼Ӧ����Ӧ�����ɣ�������survivin���������������Ⱦɫ���ƶ������﹦���ϰ���ATG7��p21CDKN1A��p53�����Է�ʽ�����DDR������ͣ��������ı��踨�����ף���ATG5��ATG7Ҳ��ͨ�����ƴٵ�������Puma��Noxa��Bax����p53�����Է�ʽ����������ϸ��������ATG7����������FOXO1������յ����ɵȡ����������Ҳϣ����ͨ����������ѹ��Ӧ��֮�������˽�Ϊ���������ɰ���֢�����ṩһ���������ݡ�
�����ο����ף�Retrograde signaling from autophagy modulates stress responses






























����˵�������а�