 )
)
�������ߣ�Elisa Corsiero (e dot corsiero at qmul dot ac dot uk)
����Queen Mary University of London, London, England
�������ߣ��˺��� �Ϻ���ͨ��ѧũҵ������ѧԺʳƷ��ѧ�빤��ϵ
�������
��������ṹ
���������ף�Ig������������ϵͳ����Ҫ��ɲ��֣����ݽṹ���ǰ�����˫�ؽ�ɫ������1���ܹ�ͨ����ԭ���Ƭ��λ�������Խ�Ͽ�ԭ��2��ͨ��Fc�μ�������ϵͳ����ϸ���鵼���߷�Ӧ��ͼһ�������ǿɷ�Ϊ���¡����-���ֿ�����ʶ��ԭ�����λ�͵���¡���壨mAbs���������Ǹ߶������ԵĿ��壬ֻ��ʶ��ԭ��ijһ�ض���λ��������40��ǰ���״η��֡��ڹ�ȥ����Щ��������Ѿ���Ϊ�����о�����֮�������㷺Ӧ���������黯����ʽϸ��������ӡ������ؼ����� [1] �� �������⣬�������20�������Ҳ��Ϊ��֢���Ƶ���Ҫ��ɲ��֡����ǵ�ҽ��Ӧ�ÿ�����չ��������֢��������ֲ��Ⱦ�� ������HIV��������������HIV������Ч������������ĿǰΪֹ������Լ30�е�����FDA��Ȩ�������ϼ��������� [2] ����ˣ����Ƕ��������Ϳ������Ȥ������������������о�Ҳ�������ӡ�
����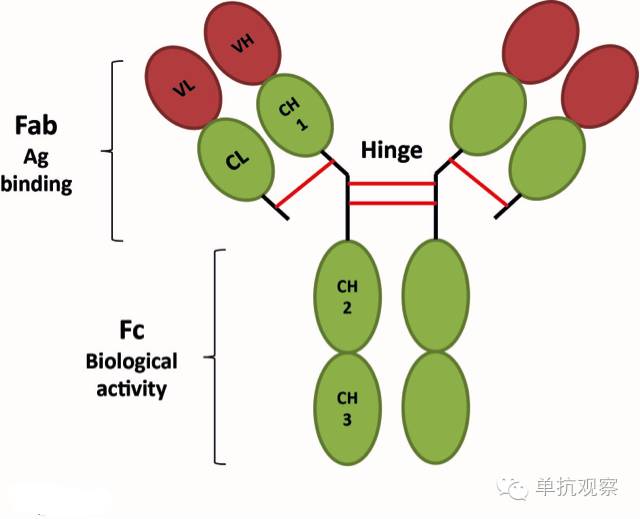
����ͼ 1. ������IgG�ṹ�� IgG���ӵļ�ʾ��ʾ��������ԭ���λ�㣨Fabs�Σ�������Fc�Σ���
��������Ŀ�����Ӱ���������ͬ��������������ͬ��������ͨ��������������ӡ�ÿ��������һ���ɱ�����VH��VL�����ڿ�ԭʶ�𣬺�һ���㶨������ЧӦ���ܣ�CH1-3��CL����ͼһ��. ����Ҳ����һ��������������Fc���֣�����ɱ�����Ϊ���֣�IgG�� IgM�� IgD�� IgA����IgE���������������ڰ�������һ����110����������ɵĿɱ�������������������Ϊ����������CDR1, CDR2��CDR3���ĸ߱���������ֲ�����ĸ����صĹǼ�����FRs����CDR3ͨ���������������Ϊ��ԭ���λ������ġ�
����������Դ�����IJ���
������Դ������ͨ���ӽ���ϸ����չʾ�������ɾ���չʾ���͵��ּ����Bϸ�������м��������ӵļ����ڱ����в�չ������������ [3] :
������С���ӽ���ϸ����CDR����ֲ����Դ�������ɱ����������Ǽ�����
�������ñ�����ԴIg�����ת����С���ڴ�ͳ���ӽ���������������Դ������
�����ӽ�������
����
�����ӽ���������һ�ֹ��ϵķ��������������Ʊ�������Ŀǰ����ʵ���������ø÷������÷��������������Bϸ���������ϸ��ϵ�ںϣ��ӽ�����Ȼ����ѡ��������������ֻ�������ض��������ں�ϸ���ܹ���ѡ���������д�
����������ι����ģ�
����Bϸ���ӱ��ض���ԭ���ߵ�С��Ƣ���з��������Bϸ������Ըÿ�ԭ�����ض�������ͨ��������ÿ�ԭ�����Ե�Bϸ����Ⱥ������ЩBϸ����ɸѡ�Ĺ�����Bϸ��ϵ�ںϣ���ͨ����ѧ���ʻ��߲����鵼�����ںϣ����ù�����ϸ��ȱ�ٱ���λ�����-������-�������ת��ø��HGPRT���Ļ������������ܲ������塣��һѡ���������Ҫ�ģ����������͵�Bϸ�������뺬�дλ�����-�װ����-������ऺ��յ�ѡ����������HAT����������������ֻ���ӽ���ϸ���ܹ�����Ϊ������ϸ���ڸ��������������ѣ����ܲ����ض������Bϸ���ڷ��Ѽ�����������ʡ����ֻ���ӽ���ϸ���ܹ����Ѻ��ơ�����ɸѡ�ܹ�����Ŀ�ĵ������ӽ�����¡����Щ�������ܹ������������з�����������ܸ÷�����Ȼ�ǵ����Ʊ��ġ������������һЩȱ�ݣ���������ֳ���ںϵ�Ч��ͨ���ܵͣ�����Bϸ���ij���״̬Ҳ����Ҫ [4] ��Ϊ�˿˷���һȱ�ݣ���2003��Michel Nussenzweigʵ���ҿ���������Ч��¡�ͱ��ﵥ���ķ���������ʽϸ�����뵥��Bϸ����ʼ���������Ķ��䣩��
������ϸ��ɸѡ����
����
�����Ʊ���Դ��������Ч�����ǻ�����ʽϸ���ǵ�Bϸ����ϸ���ּ�Ig��������������������PCR�������Լ��������⿹����������¡��
����������?
���������˵����ĵ���Bϸ���ּ�IJ����ǻ��ڴӵ���ϸ��cDNA������IgH��Ig�ʺ�Ig�˵�PCR���������ض����������ͷ���������PCR�ֱ������õ��������������ʺͦˣ���������PCR�����¡���ض����������б������������������������תȾ���˵�ϸ��ϵ����������Դ������ͨ���������������õ����塣��ϸ����ɲο���ƪ���� [4] ��
�����ɾ���չʾ�Ŀ�
����
�����ɾ���չʾ��30��ǰ���֣�����������MRC��������ѧʵ���ҵ�Greg Winter���˺�Scripps�о�������Richard Lerner, Carlos Barbas���˷ֱ������ġ��ɾ���չʾ�Ŀ��Ʊ��ĵ���������Fabs��scFvs �������ɱ�Ƭ�Σ���sdAbs �������壩 ��ͼ1��2a���� ��������ͨ�������III�¿ǵ����ں� [5-7] ��
����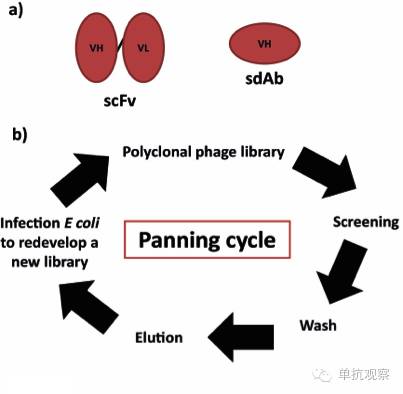
����ͼ 2. ��a�� scFv �ṹ���� ���������ṹ��һ�������ɱ�����VH����һ�������ɱ�����VL��ͨ��һ�����Խ�ͷ���ӣ� sdAb ���ң��ṹ����һ����һ�ṹ��ͨ���������ɱ����� ��b�� �ɾ���չʾ��һ�㲽�衣
����������?
�������п����ɾ���չʾ�Ŀ��ԭ���ǻ��ڽ�����������Ժ����ԣ����ͣ����ɾ���������У������ͣ����е��������ӡ�����ǿ��ٿ�ԭ�����Կ��������ɸѡ����ͼ2b�ܽᣬ������Ķ��¡�ɾ���������鿹����̶��ڴ��顢�۱���ϩ���棬��ϸ������İп�ԭ���н�ϡ��������ֶԷǽ�ϵ��ɾ����ϸ�ϴ�Ѻ�ԭ��ϵ��ɾ���ͨ��pHֵ�ı��ø������ϴ��������ɸѡ�������ɾ��屻���ڸ�Ⱦ�˾������Ʊ��µ��Ŀ�������һ�ֵ�ɸѡ�������ظ���һ��ȫѭ����ֱ�������õ�����¡���ɾ��塣�������б���ԭ�������ڱ������鿹�� [6] ��
�����ճ�ʹ�õĿ������кͽṹ������Դ
�����������е���Ҫ��Դ��The ImmunoGenetics Database ��IMGT�� ��https://www.imgt.org/���� IMGT��������ȫ����Դ���������ڸ��£����������������ϵ�������С��ڶ������õĿ���������վ��IgBLAST ��https://www.ncbi.nlm.nih.gov/igblast/��������NCBI���������Եİ��������Ŀɱ������С����ڿ���ṹ���ݿ⣬��һЩ���õ���Դ��Protein Data Bank ��PDB�� ��https://www.rcsb.org/pdb/home/home.do���� CATH���ݿ���վ��https://www.cathdb.info���� ���徧��ṹ��SACS���� IMGT-3D�ṹ-DB���м������õĹ��߿����ڿ��������о��������У�����ģ����Rosetta Online Server ��https://rosie.rosettacommons.org����3D�ṹԤ��Ŀ�����վ���ǻ���Ԥ�⣬NetNGlyc1.0 Server ��https://www.cbs.dtu.dk/services/NetNGlyc��������Ԥ����Դ����N-�ǻ���λ��Ĵ��ڣ�NetOGlyc4.0 ��https://www.cbs.dtu.dk/services/NetOGlyc�� ������O-glycosylation�ǻ���λ���Ԥ�⡣
�����������������
����
����HEK293��CHOϸ��ϵ
�������������������������������ϵͳ�еı��ѡ������ϸ��ϵ����תȾ�Ǻܹؼ��ģ��ڿ�ʼ������Ŀǰ��Ҫ����һ�¡���ȷ�������ı����ϵͳ�ܹ�Ӱ����Ҫ�Ʊ��Ŀ�����ԡ�ѡ������ϵͳ����Ҫ����Ҫȡ���ڵ����ʵķ�������Ρ��ر��ǣ����ڿ�����˵�ǻ���������ķ�������Σ��ܹ�Ӱ�쵰���۵����ȶ��ԡ��ܽ�ȡ������Ժ�����ԭ�Ե� [8] ������ϵͳ����ϸ������ĸ������ϸ�������桢ֲ���Լ��������ϵͳ��ͨ�������鶯��ϸ�������ڵ����Ʊ�����Ϊ��������ȷ��װ�����뷭������Σ�Handbook of Therapeutic Antibodies 2E ��2014������ �м��ֲ��鶯��ϸ������Ϊ����ϵͳ��Ȼ��ÿ��ϵͳ���ǻ�������������ͬ��ͨ�������о�ʵ�������ܳɹ����ڱ�����Ϊ�й������ѳ�ϸ����CHO��������̥��ϸ����HEK293��. Ȼ����CHOϵͳ��˲ʱתȾЧ�ʵͣ������ĵ��ײ���Ҳ���ߡ����HEK293��Ϊ˲ʱתȾ����Ҫϸ��ϵ����������ѡ������ϸ��ϵ��Ϊ������Ҫʱ�̼�ס�Ʊ�������ǻ�����������������ѭ����������ͬ���±��ܽ��˿�����˲ʱתȾ��HEKϸ��ϵ����:
����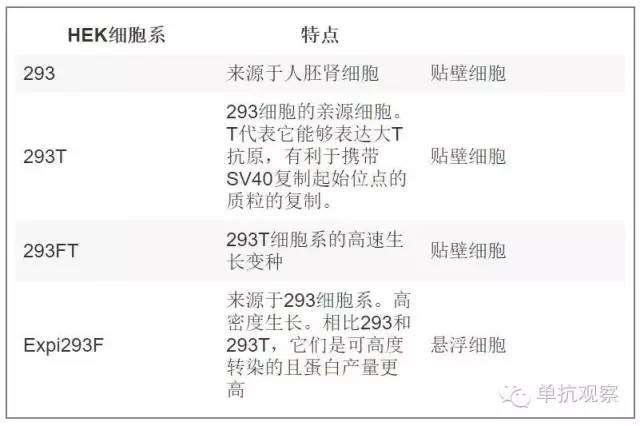
������һ�� ˲ʱתȾ��HEKϸ��ϵ��
����תȾ�Լ�
������Ʒ����תȾ�Լ��Լ��кܶࡣ��˸�������ʵ����ѡ��õ��Ǻ���Ҫ�ġ�Ҫ�ʵĵ�һ����������ϣ����õĵ��ײ�������ȡ�������øÿ�����е�ʵ������ͺʹ������������˵��Ǿ���ϩ�ǰ���PEI����PEI��һ�������Ӿۺ���ܹ���DNA�ۺϳ������ӿ���ͨ���������ý���ϸ����һ������ϸ���������ܹ���DNA�ۺ����ͷŵ�ϸ�����С�����������DNA���ܹ���ɢ��ϸ�����С�PEI�鵼��תȾ��Ҫ���������ܹ���õĵ��ײ���̫�ͣ�ͨ�����˼��𡣲������ڳ���ɸѡ������Ѿ��㹻�ˡ�����г��ϳ����˼�������ߵ��ײ�����תȾ�Լ�����Ȼ��Ǯ�������ǵ�Ч�ʸ��ߣ�������������ڶ���Ʊ�ͬһ�ֿ��壬ÿ�ζ���ʡ��һЩ�������裨���������������Ƶȣ���Thermo Fischer Scientific��վ���г�����Ҫ��תȾ�Լ���https://www.thermofisher.com�����ܽ����±���
����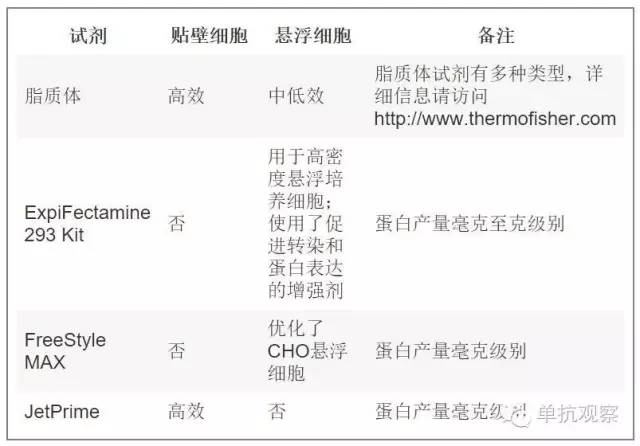
���������������Ʊ��ij���תȾ�Լ���
������������
������������Ҳ�DZ���ϵͳ����Ҫ��ɲ��֣����Ҳ���������������������Ż�����ǿ�����Ӻ���ǿ�ӵļ������Ż�������ø��ߵ�ת¼�ͷ���ˮƽ��ȷ����˵����Ҫ�õ�һЩ�ض���������/��ǿ�ӣ����ϸ��������CMV����SV40�� �ӳ����������ӣ� ����������ǿ�ӣ� ��beta-�������������ӡ����ڿ����Ʊ���˵��˲ʱתȾͨ���õ��������壬һ��������������һ�ױ���������Ȼ������תȾ��һЩȱ�ݣ�������Ҫ���������壬������ӳɱ���Ҳ��ʱ��������һЩ�о�ͨ������ͬʱ�������������������壨һ������װ���������һ���� [9] ��
��������̬����:�dz��濹��
����������������ƵĿ���Ϊ��Դ�������Ƕ�ϵ�������Щ�����Ѿ�����֤�dz��ɹ�������������һЩ�dz��濹�壬�������֯��ҩ���л��ҩ����Ч�� ����̬�Ŀ������FabƬ�Σ�������������CH1��ͨ����������ӣ��� FVƬ�Σ�V��������������-scFvs ���������Ķ�����V����������һЩ������̬�Ŀ���Ƭ�Ρ������ֲ�ͬ�����ԵĿ���Ƭ�ο����������γ�˫�����Կ����˫���塣�Էdz��濹�������̬����������������Ӧ�ý��ܵ����¿�������鿴 https://dx.doi.org/10.13070/mm.en.3.160 ��
����ʵ����ˮƽ��ͨ���Ͳ������п��崿���ķ���
����
����ͨ���Ʊ��Ŀ���ΪIgG���͵ġ���˶����ǵĴ��������ǻ��ڵ���A��G���Ͳ�����������Դ�����ܽ��������:
����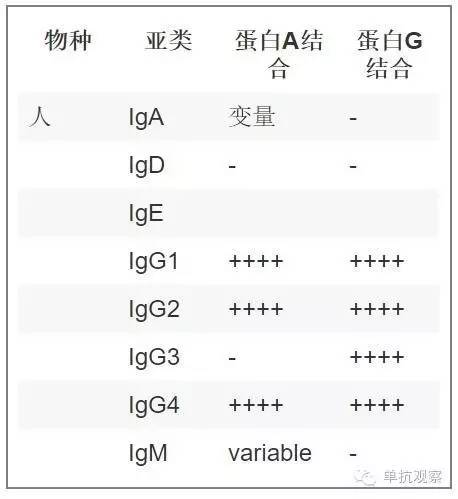
���������� �������༰��Ӧ����A/G���ˮƽ������Affinity Chromatography Handbook ��GE Healthcare��վ��
����ͨ���Ͳ������������ǻ��ڿ����Fc����̶��ڲ��������ϵ��ض����壨������A/G���Ŀ��淴Ӧʵ�ֵġ�����A��G��������ϸ�����ֱ����Խ��ɫ�������������������������Ϻ�ϽΣ�����ͨ���ı�ϴ�ѽε�pHֵ��ת��Ӧ���Ͳ���������һ���ܴӴ��ģ��������������壩��һ������Ч���տ���ķ�����GE Healthcare��վ�Ŀ������˵��������ã�����������Ʒ���Ϳ��崿������ò��裬������Ϊ�������п��崿���IJο��飨www.gelifesciences.com/Handbooks/Antibody_purification_handbook����
����С��ģ����
��������С��ģ�����ڿ���ɸѡ�к����á�����ͨ�����²���ʵ�֣� i�� ���ܹ��õ���A��G�����IJ�������Ԥ�����IJ����� ����һ���Ծ۱�ϩ������ ii�� ����A��G������96�װ����ڸ�ͨ��ɸѡ���ڵڶ���ϵͳ�У���������������У���ϴ��ϴ�ѿ�ͨ�����Ļ��������ϵͳʵ�֡�
�������ģ�����CÄKTAϵͳ
����ͨ����ʵ����ˮƽ�������ڹ�ҵ��;Ҳ��Ҫ�Ʊ���ͬ���Ŀ��壬���˼��˼����á���������Զ�����ϵͳ�����Ӳ����ʹ��Ⱦͷdz���Ҫ�ˡ����г��ϣ��м����Զ�����ϵͳ���������GE Healthcare��ÄKTA�������������ֶ�����������С��ģ��������÷�ʱ��ʱ��ʹ����Щϵͳͨ���ܵõ��ظ��Ը��õĽ�����ֶ�������÷�ʱ��ԭ������� i����Ҫ��ø���壬 ii�� ��Ʒ�ij�ʼ��̫�� iii����Ҫ����������Ʒ���Զ�����ϵͳ��Ǯƫ��Ͷ����Щϵͳ��ʹ��һЩӦ�ñ��ʡʱʡ������ʡ��Ʒ������Ҫ��������߲�����GE Healthcare�м���ÄKTA�Ļ�������Ҫ�������г�:
����ÄKTA start
����ÄKTA prime plus
����ÄKTA xpress
����ÄKTA pure
����ÄKTA avant
����ÄKTA pilot
����ÄKTA ready
����ÄKTA process
������1��5��ÄKTA����ʵ���Ҽ��𡣴�6��8�����ڹ�ҵ�������𡣸���ϸ������ģ� www.gelifesciences.com/Handbooks/Antibody_purification_handbook��
��������������干��Biacore
����Biacoreϵͳ��ͼ3a�����ñ���������干��SPR��ԭ��ʵʱ�����Ӽ�����á��÷���������������õķ��ӣ�һ������ڴ����������壩����һ���ڽ�ͨ���������Һ��������С��������������Ҳ�����Ƿ������������������Ϻ���������������������ȱ����ķ�Ӧ����ȷ����Ħ������Ħ�������Ũ�ȡ�Biacoreϵͳ���о������зdz����ã���Ϊ���ܷ�ӳ:
���������뿹ԭ����õ������ԣ�
���������뿹ԭ����õĶ���ѧ��������
�����ض����ӵ�Ũ�ȡ�
����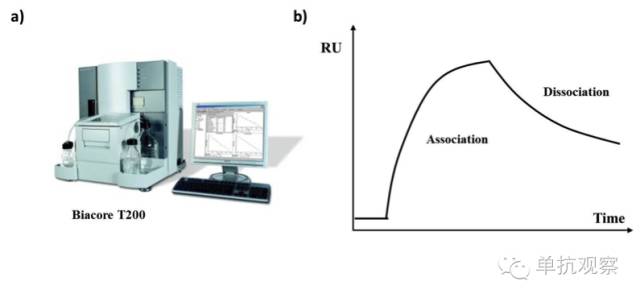
����ͼ 3. ��a�����ﴫ�������ӣ�b����Ӧͼʾ��ͼ��RU = ����Ԫ
��������Ԫ��RU���ɷ�ӳ�����źŵĸı䣬ͨ����ʱ�����Ӧ���Ը�Ӧͼ����ʽ���֣�ͼ3b����
�����ܽ �Ʊ���ҵˮƽ�Ŀ���
����
����������Ʊ��Ѿ�������������ʵ�����У���һЩҽ����;�Ŀ���Ҳ���Ʊ������ˡ���ˣ�Ӧ���ڱ�ʵ���ҵı�������Ҫ���ڿ���ɸѡ������ҽ����;��ȷʵ�����ģ�Ʊ������˵Ŀ�����Ҫ���Ӿ�ȷ�IJ��ԺͿ��Ʋ��裬��Щ����Ҫ�������ƶ��ߵζȿ���������ƽ̨��ȷ����˵����������i�������ȶ�תȾ������˲ʱתȾ��ii�������ﷴӦ������ϸ����iii���ӷ��ڲ�����ȥ��ϸ�����Լ����ö��ֲ������˼�����ϸ�����������д������壬iv���ı仺��Һ��������������ϣ�����䷽�������ٴ���;��������Щ���趼��Ҧ���ϵ��������Ʋ�����ȷ����ȫ��������ԡ����������Щ���趼ֻ���ڹ�ҵˮƽ�����DZ�ʵ����ˮƽʵ�֡�
���������
����
����Weiner G. Building better monoclonal antibody-based therapeutics. Nat Rev Cancer. 2015;15:361-70
����Li J, Zhu Z. Research and development of next generation of antibody-based therapeutics. Acta Pharmacol Sin. 2010;31:1198-207
����Voula Kodoyianni MH, Marjorie Curet, Rachel Kravitz, Brian Schram A Novel Process for Developing Fully Human Monoclonal Antibodies. 2012.
����Wardemann H, Kofer J. Expression cloning of human B cell immunoglobulins. Methods Mol Biol. 2013;971:93-111
����Arbabi-Ghahroudi M, MacKenzie R, Tanha J. Selection of non-aggregating VH binders from synthetic VH phage-display libraries. Methods Mol Biol. 2009;525:187-216, xiii
����Chan C, Lim A, Macary P, Hanson B. The role of phage display in therapeutic antibody discovery. Int Immunol. 2014;26:649-57
����Hoogenboom H. Selecting and screening recombinant antibody libraries. Nat Biotechnol. 2005;23:1105-16
����Krista Steger PD, James Brady, Ph.D., Meg Duskin, Karen Donato, Ph.D. Literature Review: CHO versus HEK Cell Glycosylation. wwwmaxcytecom.
����Dodev T, Karagiannis P, Gilbert A, Josephs D, Bowen H, James L, et al. A tool kit for rapid cloning and expression of recombinant antibodies. Sci Rep. 2014;4:5885
����Doerner A, Rhiel L, Zielonka S, Kolmar H. Therapeutic antibody engineering by high efficiency cell screening. FEBS Lett. 2014;588:278-87
���������ɵ����۲�������������






























����˵�������а�